A Look At Some Abilities Which Are Similar In Humans And Nonhumans (1986)
©1986, 2013 by Dallas Denny
Source: Dallas Denny. (1986, 26 April). A look at some abilities which are similar and humans and nonhumans. Paper for Dr. Jon Kaas, Psychology 344, George Peabody College of Vanderbilt University.
A Look At Some Abilities Which Are Similar In Humans And Nonhumans
By Dallas Denny
For Dr. Jon Kaas
Psychology 344
April 26, 1986
Abstract
Abstract
Homo sapiens shares with other mammals many basic aspects of brain functioning. While our brains of course have specific adaptations which reflect our cognitive and language abilities, the basic mammalian plan is present, and learning, memory, and perhaps cognition are only quantitatively different from other mammals. Many of our abilities may be relatively independent of our much-touted intelligence. This paper will present a discussion of cognitive and language abilities in non-human animals, and will conclude with some speculations about some areas of functioning which do not require high intelligence, yet which are quite complex.
Alfred Russel Wallace, the co-discoverer (with Sir Charles Darwin) of natural selection, believed firmly that organisms are in all regards perfectly adapted to their environments (Brackman, 1980; Gould, 1977, 1980; Wallace, 1890). Although biologists now know this is not so (Gould, 1980), it is clear that natural selection is the major mechanism which leads to diversity of species, and that many aspects of morphology and behavior have evolved because they increase the probability of survival of genetic material (Alcock, 1979). Lorenz (1973) noted, “As natural selection ‘breeds’ structures which fulfill particularly well a certain survival function, they finally look as though they had been created for the very purpose by a wise and benevolent spirit” (p. 22). Lorenz (1965) has elsewhere pointed out that this is as true for behaviors as it is for morphology. In our species, the morphological trait which has perhaps attracted the most attention and generated the most heat and the least light is the brain.
Much of the interest in the brain has come about because of its function rather than its form: If the brain is responsible for the wondrous complexity of human behavior, for our cognitive and language abilities, for the creation of Macbeth and the Mona Lisa and NASA and the pun, then surely it must itself be a wondrous creation. A history of the study of the human brain is beyond the scope of this paper, but I would maintain that through the years many of the questions asked about the brain and many of the interpretations of data gathered about the brain reflect our chauvinistic attitudes about ourselves as a species. A number of instances of this were discussed in detail by Gould (1981), who revealed how conscious and unconscious bias can affect the work of even the most conscientious scientist.
The fact is that except for a rather well-developed cerebral cortex and perhaps more lateralization of function in the cerebral hemispheres than is found in other mammals, the human brain is rather unremarkable as an organ (Geschwind, 1979). The basic parts of the human brain are in the general mammalian plan, and we might therefore expect that many human abilities are representative of mammals and not particularly special at all.
Few remarkable insights about human nature have emerged from the study of the brain itself. It is only when (1) structure and function are considered together (Welker, 1975) or (2) the comparative method (Alcock, 1980) is used, that is, when the brain is viewed in relation to the brains of other primates, living and extinct, that the most useful information about human nature has been obtained. For example, the fossil record clearly illustrates the brain size of ancestral hominids increased dramatically during the past five or six million years (Jerison, 1977; Weaver, 1985); this suggests there was strong selective pressure for larger brains in our remote ancestors. Another example concerns two small regions of the cerebral cortex called Broca’s and Wernicke’s areas. These areas are important for speech in humans (Kolb & Whishaw, 1980); their presence or absence in ancestral hominids and in living primates can tell us much about the antiquity of speech. These and similar avenues of research can tell us much about ourselves, and leave a legacy for future generations; compare them with the fruitless search of Karl Lashley for the locus of memory (Lashley, 1950) or the almost forgotten work of the phrenologists of the last century (Gould, 1981).

Dr. Roger Fouts with Washoe, 1972
The general high opinion of the intellectual capacities of our species has led to artificial distinctions between ourselves and other animals. One area in which humans were long thought to differ from other animals is in the use of symbolic communication. Humans were held to be the only animals to use language. During the past few decades, this has been challenged on a number of fronts. The first definitive evidence of language in non-human animals was the Nobel-winning work of Karl von Frisch (1953, 1967, 1974), who found bees use a symbolic “waggle” dance to inform other bees of the whereabouts of nectar-bearing flowers. And in 1969 Gardner and Gardner published the results of an attempt to teach the chimpanzee Washoe to communicate by use of American Sign Language. A number of other apes have been taught to communicate by use of sign language, manipulation of abstract plastic symbols, and by pressing buttons attached to a computer (see Linden, 1974 for a review of early studies). To the casual observer, these apes indeed seem to be communicating; however, the Gardners’ work with Washoe has been criticized on methodological grounds, and linguists and psychologists have advanced subtle arguments which indicate that the apes are not using language (Premack, 1979; Terrace, 1979, 1985). To me, they seem to perhaps be nitpicking. I have trouble following some of the arguments; I am reminded of the comment of one of Broca’s followers that “I have noticed for a long time that, in general, those who deny the intellectual importance of the brain’s volume have small heads” (Gould, 1980, p. 146). Recent work has extended some of the findings with apes to other animals, notably the African grey parrot (Pepperberg, 1985) and California sea lions (Schusterman, 1985).
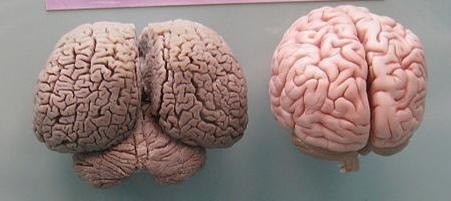
The brain of a dolphin is on the left. A human brain is on the right.
Closely related to language is the ability or group of abilities which we call intelligence. Unfortunately, we aren’t sure just what comprises intelligence in humans (cf Sattler, 1974), so we aren’t in a good position to define it in animals. Certainly there are reports of the cognitive abilities (and lack of cognitive abilities) in animals. Such reports can be deceptive, as noted by Barash (1982), who has pointed out that operant conditioning and social learning studies usually don’t take into account the natural history and behavior of the species. Barash gives as an example detour problems. Squirrels, which are considered to be less intelligent than dogs and which have simpler brains, do much better at tasks which require them to initially go away from a goal in order to eventually get to it. Barash explains that squirrels, which are arboreal animals, must routinely go away from food or conspecifics in order to ultimately approach them. Dogs, in their evolutionary history, have rarely had to avoid objects which they wanted to approach, and consequently perform worse than squirrels; however, dogs are superior to squirrels in other tasks.
Ethologists know many behaviors which are quite complex and which seem to require considerable intelligence may be relatively inflexible behaviors which are elicited by specific stimuli (Tinbergen, 1951). Many “intelligent” behaviors of animals are little more than highly specific innate responses to specific environmental stimuli. There is nonetheless evidence of insight learning and concept formation in nonhuman mammals. Thorndike (1911) was convinced cats were very stupid because they took a long time to escape from puzzle boxes. Donald Griffin (1976, 1978), on the other hand, is convinced that the development of a cognitive ethology is possible, and suggests a number of needed areas of investigation.
Kohler (1925) found chimpanzees were much better than dogs at going around a barrier which separated them from food. And Tolman (1948) found rats find their way about by the development of “cognitive maps,” that is, they were able to find their way about in his test apparatus without specific cues from the environment if they were allowed to experience the relationship between those cues.
These findings don’t provide conclusive proof that animals have cognition. A number of other findings, however, provide stronger evidence. Herrenstein, Loveland, & Cable (1976) demonstrated that pigeons in operant conditioning chambers are able to discriminate photographs of man-made from natural objects. The pigeons were highly accurate and were able to make subtle discriminations. Kohler (1925) reported that chimpanzees stacked boxes which were in their cage and stood on them in order to reach fruits which were suspended from the ceiling. The chimpanzees would also use sticks to pull food into their cage. Sultan, an adult male, fastened several sticks together in order to reach food. Jane Van Lawick-Goodall (1968, 1971) noted that free-living chimpanzees use objects as tools. They crush leaves and use them to soak up water for drinking. They also select twigs, strip the bark away, and stick them into holes in termite mounds. When they withdraw the sticks, termite soldiers are clinging to them, and they eat the termites. The chimpanzees appear to carefully select the stick and may reject many before finding just the right one. Chimpanzees have also been observed to hurl rocks and sticks at enemies.

Chimpanzee Fishing for Termintes
Chimpanzees are apparently aware of themselves. Gallup (1970) anesthetized adult animals; while they were unconscious, he painted their ear or a spot on their forehead with red dye. When the animal awoke, it was allowed to see itself in a mirror. The chimpanzees, upon seeing their images, reached for and touched the dyed spots. Control animals, which were anesthetized but not painted, did not do so. Lower primates and other mammals don’t do this. Rumbaugh (1985) reported that he had placed a closed circuit television camera and monitor in a chimpanzee’s cage. The chimp spent a good deal of time posing in front of the camera while it watched the monitor. The chimpanzee then stood on a chair in front of the camera and opened its mouth, while it cut its eyes toward the monitor. It was, of course, too dark to see down its throat. The chimp then fetched a flashlight, got back up on the chair, shone the flashlight down its throat, and looked down its throat by watching the monitor.
Animals often act as if they are able to predict the future behavior of other animals. They also engage in behaviors which provide other animals with information about their probably future behavior. Ethologists have long appreciated the communicative value of displays and other types of signals. These signals can be conveyed by chemical, visual, auditory, tactile, or even electrical means (Alcock, 1979). Displays are often highly species-specific, may be rigidly ritualized and stereotyped, and may communicate only limited information. However, for a display to be effective, information must be conveyed to another animal which must in turn process that information and alter the probability of at least some of its behaviors. In some species displays are used for little more than attracting mates or fooling predators (Wickler, 1968), but highly social animals have elaborate communication systems. Vervet monkeys, for example, have a system of grunts which can convey information not only that a predator is present, but that a predator of a particular type is present (Cheney, 1984). Furthermore, vervet monkeys are able to distinguish alarm cries of members of their group from cries by members of an out-group (Cheney, 1984).
The nuances of such advanced communication systems in other species are difficult to convey in print. However, nonverbal communication in humans has been studied extensively since Darwin’s (1872) Expression of the Emotions in Man and Animals. Even a casual perusal of Desmond Morris’ Manwatching shows the varied and often subtle ways in which humans use nonverbal signals to communicate intent and how we as individuals receive and use those signals to make predictions about the future behavior of others. Few humans from any culture would have any trouble guessing the mood of a red-faced man who was rapidly approaching with clenched fists raised in the air. The eyebrow flash (Eibl-Eibesfeldt, 1975) is a subtle greeting found in all cultures. People in all cultures respond to it, but no one in any culture was aware it even existed before Eibl-Eibesfeldt described it.

Eyebrow Flash Time Lapse
See the above eyebrow flash in motion here.
The ability of non-human animals to predict the future behavior of other animals implies some sort of inner schema. I think this schema is sufficiently different from cognition to warrant its own descriptive term. I know of no better word than awareness, a much-maligned word that is synonymous with consciousness.
I have come into this line of reasoning by a circuitous route which has included observations of my dog and observations of mentally retarded individuals. Severely and profoundly retarded persons (and my dog) lack that ability which we call cognition or intelligence. Yet my dog consistently monitored my movements and tone of voice and made accurate predictions about my behavior. For example, I could look at her and raise my eyebrows, and she should would begin her “go outside” dance. Of course, many persons have described the behavior of their pets, with little impact (since Lloyd Morgan in 1894), but what of retarded persons?
I once analyzed the skills necessary for successful food-stealing, at which institutionalized mentally retarded persons are often quite accomplished. I was astounded by what is required to successfully steal food. First, it is necessary to locate the food in the environment. Second, it is necessary to make some judgement about the desirability of the food. Third, it is necessary to make some judgement about the probability of retaliation from the person who owns the food. Fourth, it is necessary to make some judgement about the probability of retaliation by staff. Fifth, it is necessary to optimize the time of stealing so there is a high probability of getting the food in the mouth and swallowed before retaliation occurs. That requires monitoring the attention of other individuals in the environment and making some judgement about when they are not paying attention. And yet, mentally retarded persons are surprisingly good at stealing food. And so are non-human animals.
I have not yet done a formal analysis of food-stealing in mentally retarded persons. Perhaps they are measurably less proficient at stealing food than non-retarded persons. What I find important is that notwithstanding their lack of intelligence, these individuals have the perform so complicated a task quite well. I believe the reason these individuals do so well at is that we (humans) share with other mammals a basic mechanism of brain functioning which makes us similar in terms of our ability to learn, remember, and determine intent of other animals, and these abilities are independent of what has been called intelligence.
There are many areas I have not reviewed here, and interesting findings from a number of fields which I hope to one day use in conjunction with data from experiments with mentally retarded persons to more forcefully make my point. In particular, a discussion of the importance of operant conditioning, social learning, memory, and cultural transmission of information in man and animals would seem to be necessary to argue convincingly about basic similarities of brain functioning in man and animals.
References
Alcock, J. (1979). Animal behavior: An evolutionary approach (2nd edition). Sunderland, Massachusetts: Sinauer Associates, Inc.
Barash, D.P. (1982). Sociobiology and behavior (Second ed.). New York: Elsevier.
Brackman, A.C. (1980). A delicate arrangement: The strange case of Charles Darwin and Alfred Russel Wallace. New York: Times Books.
Cheney, D.L. (1984). Category formation in vervet monkeys. In Harre, P.; & Reynolds, V. (Eds.), The meaning of primate signals. Cambridge: Cambridge University Press.
Darwin, C. (1872). Expression of the emotions in man and animals. London: Murray (Reprinted 1965, by University of Chicago Press, with Preface by Konrad Lorenz).
Eibl-Eibesfeldt, I. (1975). Ethology: The biology of behavior (2nd Ed.) New York: Holt, Rinehart, and Winston.
Gardner, B.T.; & Gardner, R.A. (1969). Teaching sign language to a chimpanzee. Science, 165, pp. 664-672,
Geschwind, N. (1979). Specializations of the human brain. In The Brain. San Francisco: W.H. Freeman and Company.
Griffin, D.R. (1976). The question of animal awareness: Evolutionary continuity of mental experience. New York: The Rockefeller University Press.
Griffin, D.R. (1982). Prospects for a cognitive ethology. The Behavioral and Brain Sciences, pp. 527-538.
Gould, S.J. (1977). Ever since Darwin. New York: W.W. Norton and Company.
Gould, S.J. (1980). The panda’s thumb: More reflections in natural history. New York: W,W. Norton and Company.
Gould, S.J. (1981). The mismeasure of man. New York: W.W. Norton and Company.
Herrenstein, R.; Loveland, D.H.,; & Cable, C. (1976). Natural concepts in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 2, pp. 285-302.
Jerison, H,J. (1977). Evolution of the brain. In Wittrock, M,C. (Ed.): The human brain. Englewood Cliffs, NJ: Prentice-Hall, Inc.
Kolb, B.; & Whishaw, I.Q. (1980). Fundamentals of human neuropsychology. San Francisco: W.H. Freeman and Company.
Kohler, W. (1925). The mentality of apes. New York: Liveright.
Lashley, K.S. (1950). In search of the engram. In: Symposia of the Society for Experimental Biology IV, Physiological mechanisms in animal behaviour. Cambridge: Cambridge University Press.
Linden, F. (1974). Apes, men, and language. New York: Penguin Books.
Lorenz, K. (1965). In Darwin, C. (Ed.): The expression of the emotions in man and animals. Chicago: University of Chicago Press.
Lorenz, K. (1980). Behind the mirror. New York: Harcourt Brace Jovanovich.
Morgan, C.L. (1894). An introduction to comparative psychology. London: Scott.
Morris, D. (1977). Manwatching: A field guide to human behavior. New York: Harry N. Abrams, Inc.
Pepperberg, I. (1985), Evidence for conceptual and verbal abilities in the African grey parrot: Labeling of cardinal sets and categorical discriminations. Presented at the eleventh annual convention of the Association for Behavior Analysis, Columbus, Ohio, 24-27 May.
Premack, D. (1979). Species of intelligence. Debate between Premack and Chomsky. The Sciences, 19, pp. 6-23,
Rumbaugh, D. (1985). Presented at the annual meeting of the Animal Behavior Society, Durham, NC, 6-11 June.
Sattler, J.M. (1974). Assessment of children’s intelligence (Revised reprint). Philadelphia, PA: W.B. Saunders Company.
Schusterman, R.J. (1985). Semantic comprehension in California sea lions. Presented at the annual meeting of the Animal Behavior Society, Durham, NC, 6-11 June.
Terrace, H.S. (1979). Is problem solving language? A review of Premack’s “Intelligence in apes and man.” Journal of the Experimental Analysis of Behavior, 31, pp. 161-175.
Terrace, H.S. (1985). In the beginning was the “name.” American Psychologist, 40(9), pp. 1011-1028.
Thorndike, E.L. (1911). Animal intelligence. New York: Macmillan.
Tinbergen, N. (1951). The study of instinct. London: Oxford University Press.
Tolman, E.C. (1948). Cognitive maps in rats and men. Psychological Review, 55, pp. 189-208.
van Lawick-Goodall, J. (1968). Behavior of free-living chimpanzees of the Gombe Stream area. Animal Behavior Monographs, 1, pp. 165-311.
van Lawick-Goodall, J. (1971). In the shadow of man. Boston: Houghton Mifflin.
von Frisch, K. (1953). The dancing bees. New York: Harcourt Brace Jovanovich.
von Frisch, K. (1967). The dance language and orientation of bees. Cambridge, Massachusetts: Harvard University Press.
von Frisch, K. (1974). Decoding the language of the bee. Scientific American, 185, pp. 663-668.
Wallace, A.R. (1890), Darwinism. London: Murray.
Weaver, K.F. (1985). The search for our ancestors. National Geographic, 168(5), pp. 560623.
Welker, W. (1972). Brain evolution in mammals: A review of concepts, problems, and methods, pp. 251-344.
Wickler, W. (1968). Mimicry in plants and animals. New York: McGraw Hill Book Company.